قصتنا مع الشمبانزي
حدثَت هذه الواقعة في جامعة أكسفورد، يوم السبت
٣٠ يونيو ١٨٦٠م، بعد ستة أشهرٍ مِن صدور كتاب «أصل
الأنواع» لتشارلس داروين C.
Darwin. انعقد اجتماع المؤتمر
السنوي للجمعية البريطانية لتقدُّم العلوم في هذا
التاريخ، وحضره نحو ٧٠٠ شخص، وكان الموضوع هو
«التطوُّر.»
بدأ الاجتماع بمحاضرة للدكتور «جون و. دريبر»
J. W. Draper
الأمريكي، وكانت عن «التطوُّر الفكري لأوروبا،
بالإشارة إلى أفكار المستر داروين». استغرقَت
المحاضرة ساعةً، ثم بدأ غَيْرُه يتحدثون، يعالجون
نفسَ القضية، على المنصة كان يجلس عددٌ من رجال
الكنيسة، من بينهم الأُسْقُف «صمويل ويلبرفورس»
S.
Wilberforce. عُرِفَ عن هذا
الرجل أنه مُحاوِرٌ بارع. طَلَب الجمهورُ منه أن
يتحدث، فحاول التملُّص. لم يكن الرجلُ متعمقًا في
العلوم البيولوجية، ولم يكن على ما يبدو قد قرأ
كتابَ داروين بالعناية الكافية. كل ما كان يعرفُه
قد جاء عن جُلوسِه واستماعِه إلى السير «ريتشارد
أوين» R. Owen،
عالِم التشريح الشهير، وأحد كبار معارضي داروين،
عندما وقَف «ويلبرفورس» ليتحدث، انتقد النظرية
بشكلٍ ساخرٍ للغاية، وجعلها تبدو سخيفةً مضحكة.
وفي نهاية حديثه التَفَتَ إلى الشاب توماس هنري
هَكْسلي T. H.
Huxley، وكان يجلسُ بجواره
على المنصة، وسأله في تهكُّم:
«تقولُ إنك من سُلالة القرود، فهل يا
تُرى وصلَكَ منها الإرثُ عن طريق جَدَّتك
أم عن طريق جَدِّك؟»
هنا ضرب «هَكْسلي» بيده على ركبتَيه، وتَمتَم
لنفسه: «لقد سَعَيْتَ إلى حَتْفِك بظِلفِك. لن
أتركك!»
جلس الأُسْقُف وعاصفةٌ من التصفيق والضحك تملأ
القاعة. طلَب الحضور من «هَكْسلي» أن يتكلم.
نَهَضَ متحديًا. بدا وكأنه قد تجاهَلَ أنَّ
الأُسْقُفَ لم يكن يُهاجِم فكرة التطور، وإنما
يهاجمه هو شخصيًّا. شرح «هكسلي» الأفكارَ
الأساسيةَ للنظرية، وفضَح ما رأى أنه جهلٌ في حديث
«ويلبرفورس». وفجأةً تحوَّل ليواجه الأُسْقُف
المغرور. تَفَحَصَّهُ بعينَيه أولًا، كما لو كان
أحفورةً قديمةً غريبةً، ثم قال:
«إنني لا أخجل من أن يكون القردُ من
أسلافي، لكنني أخجلُ من أن أرتبط برجلٍ
يستغلُّ مواهبَه وفَصَاحَتَهُ في أن
يُعَهِّرَ الحقيقة ويُخفيَها، ويُشَوِّهَ
صورةَ علماء أفنَوا حياتَهم يبحثون
عنها!»
كانت هذه هي المعركة الأولى في حربٍ طويلةٍ
مستمرة — ما زالت — حتى اليوم. القضية لم تَمُتْ،
بل إنها في الحق تزدادُ اشتعالًا، تُذكيها نتائجُ
علميةٌ جديدة، يأتي معظمها عن علم الوراثة
الجُزَيئية الحديث، الذي حَوَّلَ كثيرًا من
اهتمامه الآن إلى دراسة جينومات الرئيسات
Primates، ولا
سيَّما الشمبانزي
Chimpanzee
(أو الشمب
Chimp).
مَن يكون الشمبانزي؟
هناك من الشمبانزي نوعان يمثِّلان أرقى
القردة العليا؛ بان تراجلودايتس
Pan
troglodytes وهو
الشمبانزي الشائعُ المعروفُ ويستوطن الضفَّة
اليمنى من نهر الكونغو، وبان بانيسكَص
Pan
paniscus أو البُونُوبُو
Bonobo،
ويُسمَّى الشمبانزي القزم، أو شمبانزي الضفَّة
اليسرى لنهر الكونغو. يعتقد علماءُ البيولوجيا
أن الشمب هو أقربُ أقاربِنا في سجل التطور،
وأنه قد جاء عن انشعابٍ من سلَفٍ شائعٍ معنا
منذ ٤–٧ ملايين عام، وأنه يُشاطِرُنا في ٩٨٪
من المادة الوراثية كما قالت ماري كلير كينج
M. C.
King عام ١٩٧٥م — الأمرُ
الذي حدا بجاريد دياموند J.
Diamond أن يُطلِقَ على
البشرِ اسمَ «الشمبانزي الثالث.»
ينتشرُ الشمبانزي في الغابات الاستوائية
والسافانا المطيرة بغربِ ووسطِ أفريقيا، لكنَّ
موطنَه قد اختُزلَ كثيرًا في السنين الأخيرة،
وأصبحَ النوعُ مهددًا بالانقراض؛ إذ يبلغ
تعداده الحالي ما بين ١٠٠٠٠٠ و٢٠٠٠٠٠ فرد،
وكان منه الملايين منذ مائة سنة!
بِنيةُ الشمبانزي أقوى كثيرًا من بِنية
البشر، ويصل طول الأنثى البالغة إلى ٦٦–١٠٠سم،
أما الذكر البالغ فما بين ٩٠ و١٢٠سم. أما وزنُ
الأنثى البالغة فيتراوح ما بين ٢٦ و٥٠كجم،
والذكر البالغ ما بين ٣٥ و٧٠كجم. يُغطِّي
الجسمَ شعرٌ أسودُ داكنٌ خَشِن، فيما عدا
الوجه (إلَّا من لحيةٍ بيضاءَ قصيرةٍ في
البالغين من الذكور والإناث) وأصابع اليد
والقدم وراحة اليد وباطن القدم وتحت الإبط.
الإبهامُ وإصبعُ القدم الكبيرة يُمَكِّنان
الحيوان من القبضة المتينة بيدَيه ورجلَيه.
وهو بلا ذيلٍ وذراعاه طويلتان أطول من
رجلَيه.
يصل الذكَرُ إلى النضج الجنسي في عمر ٧-٨
سنوات، والأنثى في عمر ٦–١٠ سنوات. تبلغ فترةُ
الحمل نحو ثمانية أشهر، ويندُر أن يُولَد
توائم. تُقْطَمُ الصغار على عمر ٣ سنوات، وإن
بَقِيَت ملازمةً للأم سبع سنوات أو نحوها.
يبلغ متوسِّط طول العمر في الأَسْر نحو ٦٠
عامًا، أما في الغابة فيتراوح ما بين ٣٥ و٤٠
عامًا.
الشمبانزي حيوانٌ اجتماعي للغاية يعيش في
مجاميعَ أو مجتمعاتٍ تتراوح أعدادها ما بين ٢٠
و١٥٠ فردًا. على أنه يتحرَّك معظمَ الوقتِ في
مجاميعَ صغيرةٍ من بضعة أفراد لا أكثر (٦ أو ٧
أفراد). وهو حيوانٌ نهاري شَجَري وأرضي، يُنفق
وقتًا متساويًا في الأشجار وعلى الأرض. يمشي
الحيوان طبيعيًّا على أربع، لكنه يستطيع المشي
على اثنتَين لمسافاتٍ قصيرة. يستخدم الحيوانُ
تعبيراتِ الوجه والصوتَ ولغةَ الجسد، حتى
الأحضان والقُبَل، للتواصل مع أفراد جماعته،
بل ولقد يَسْتَجدي الفردُ الطعامَ من زميله،
فيقتربُ منه مُستَعطِفًا — مفتوح
اليدَين!
غذاءُ الشمبانزي أساسًا نباتيٌّ (الثمار،
والأزهار، والبذور، والسوق، والقلف، والدرنات)
مع بعضِ الحشراتِ والفرائسِ الصغيرة. وقد
رُصدَت حالاتٌ يقومُ فيها بالصيد، إن تكن على
ما يبدو حالاتٍ وقائية يحرِّكُها الجوع. على
أن أفرادَ الشمبانزي قد يتجمَّعون لاصطياد
قردٍ وأكلِ لحمه؛ الأمر الذي قد يعني أن
الشمبانزي قادرٌ كالبشر على الصيد الجماعي،
وأنه يَستَطعِمُ اللحمَ.
قصتنا مع الشمبانزي
في تسعينياتِ القرنِ السابعَ عشَر، قام
العالِم الإنجليزي «إدوارد تايسون»
E. Tyson
بتشريح أول شمبانزي، ولاحظ أَنَّ بين مُخه
ومُخ الإنسان تشابهًا «مدهشًا»، لكنَّ عالِمَ
التشريح السير «ريتشارد أويِن»، في القرنِ
التاسعَ عشَر، قال إن هناك بين المُخَّين
فارقًا واضحًا للغاية، وليس بين البشر مَن
يمكن اعتباره شمبانزي مُحَوَّرًا. كان أويِن
معارضًا عنيفًا لآراء داروين في التطور،
ولآراء الفرنسي «جان بابتيست لامارك»
J. B.
Lamarck مِن قبله؛ ومن
ثَم فَقَدْ بَحَث عن شيءٍ يتفرَّد به مخُّ
الإنسان ولا يُوجَد له مثيلٌ في بقية
الرئيسات. وقد وجَد ضَالَّتَه في طَيَّةٍ
صغيرةٍ تُوجَد بمؤخرِ المُخ البشري ولا تُوجَد
في غيره، واعتبرَها مَركزَ العقل والفِطنة،
حتى ليضَع بسببها الإنسانَ في مرتبةٍ رفيعةٍ
وَحْدَه، إلى أن جاء خصْمُه اللدود «توماس
هنري هكسلي»، ليَنفيَ الروايةَ ويقولَ إن
أويِن قد أهمَل في حماقةٍ وجودَ «الطيَّة
البشرية» في القِرَدة العليا.
وعلى بداية القرن العشرين، بدأ العلماء
ينتبهون إلى الرابطة البيوكيماوية بين البشر
والشمبانزي، فاكتشَفوا أن شَكْل الكثير من
بروتينات الشمب يُشبِه بروتينات البشر، بل
ويكاد يطابقُها. وعلى أوائل ثمانينيات ذلك
القرن، تحوَّل العلماء من البروتينات إلى مادة
الوراثة — واكتشفوا أن مادتَنا الوراثيةَ تشبه
المادةَ الوراثيةَ للشمب كثيرًا؛ في
الكروموزومات وفي التشريح الجُزَيئي.
الكروموزومات
يحمل الكائن الحي جهازَه الوراثيَّ في نواةِ
كلِّ خليةٍ في خلاياه. يتألفُ هذا الجهازُ
أساسًا من عددٍ «مزدوج» من الكروموزومات
Chromosomes
مُحَدِّد في كل نوع (هو في الإنسان مثلًا ٢٣
زوجًا، في الشمب ٢٤ زوجًا، وفي ذبابة الفاكهة
٤ أزواج)، يأتي نِصفُ العدد من الأب (في
الحيوان المَنَوي) والنصفُ الآخر من الأم (في
البُوَيْضة). الكروموزوماتُ أجسامٌ
عَصَوِيَّةٌ تتكوَّن أساسًا من دنا
DNA
ملفوفٍ لفًّا مضاعفًا، مع بروتيناتٍ مرافقة،
وبه منطقةٌ منقبضة تُسمَّى السنترومير
centromere
تقسم الكروموزوم إلى ذراعَين غير متساويتَين
عادة، واحدة قصيرة (ق =
p)
والأخرى طويلة (ط =
q). في
كلٍّ من طَرفَي جديلتَي كلِّ كروموزوم يُوجَد
تتابُع من القواعد يُسمَّى التيلومير
telomere
وظيفتُه حماية الجديلة أثناء الانقسام. يتآكل
التيلومير مع توالي الانقسام وقد يُعاد
تطويلُه، ويكون ذلك عن طريق إنزيم يُسمَّى
التيلوميريز، فإذا ما أصبح التيلومير قصيرًا
للغاية في الخلايا الجسدية غَدَا عُرضةً عند
الانقسام للأخطاء التي قد تؤدِّي إلى
السرطان.
إذا صُبغَت الكروموزومات في المعمل بصبغة
جيمسا
Giemsa
اتخذَت نمطًا مُعَيَّنًا من التشريط (شرائط
داكنة وفاتحة متعاقبة ذات أحجامٍ مختلفة) يمكن
به تمييزُ الكروموزومات بعضها عن بعضٍ للبشر
٢٣ زوجًا من الكروموزومات يُسمَّى كلٌّ منها
أوتوزومًا، فيما عدا اثنَين، هما كروموزومَا
الجنس، واحد يسمَّى س
X (ومنه
اثنان في الإناث وواحد في الذكور) والآخر يسمى
ص
Y (ومنه في
الذكور نسخةٌ واحدة). تُرقَّمُ كروموزومات
الإنسان حسب طولها من ١ (الأطول) حتى ٢٢،
بجانب كروموزومَي الجنس. ويُطلَق على الذراع
اسم ق أو ط حسب طوله كما ذكرنا. فإذا قلنا ٧ط
7q، فإن
هذا يعني الذراع الطويلة للكروموزوم السابع،
كما تُقسم كل ذراع إلى ٣ مناطق؛ ١، ٢، ٣،
ويبدأ الترقيم في كلٍّ من السنترومير في اتجاه
التيلومير. وعلى هذا فإن ٧ط١
7q1 تعني
المنطقة الأولى (الملاصقة للسنترومير) من
الذراع الطويلة للكروموزوم السابع. وهناك
تقسيماتٌ أدنى من ذلك، تقسم فيها كل منطقة إلى
شرائط داكنة وفاتحة تُرقَمُ هي الأخرى. وقد
يُقسم الشريط إلى تحت شرائط (انظر الشكل
١).
الطفرات الكروموزومية
كثيرًا ما يحدث أثناء عملية الانقسام
الاختزالي للخلايا أن يُعاد ترتيب مقاطع كاملة
من الكروموزومات، فتتحرك داخل نفس الكروموزوم،
أو إلى كروموزوم آخر، الأمر الذي يتسبب في
تغيرات في مورفولوجيا الكروموزوم نفسه. يُطلق
على هذه الحالات اسم الشذوذ الكروموزومي
aberrations،
ومنه صور عديدة. فالانقلاب
inversion،
يعني أن ينكسر الكروموزوم في مكانين منه، ثم
ينقلبُ هذا المقطعُ المكسور ويستقر مقلوبًا في
نفس موضعه. والاقتضابُ
deletion
يعني أَن يَفْقدَ الكروموزوم قطعةً منه،
والإيلاج
insertion
يعني أن يُضاف إلى الكروموزوم مقطع من مصدر
مجهول ويصبح بعضًا منه. أما الانتقال
translocation
فهو أن يتحرك جزء من مادة الكروموزوم إلى مكان
آخر، قد يكون بنفس الكروموزوم أو يكون في
غيره.
كروموزومات الشمبانزي متشابهة في التشريط مع
كروموزومات البشر. وأهم اختلاف هو أن للبشر
زوجًا من الكروموزومات أقل من الشمبانزي (وكل
القردة العليا)، وهناك اختلافاتٌ أخرى بين
البشر والشمبانزي؛ إذ تُوجد انقلاباتٌ في
مقاطعَ كروموزوميةٍ لكروموزومات البشرية: ١،
٤، ٥، ٩، ١٢، ١٥، ١٦، ١٧، ١٨، بجانب طَفْرة
انتقال، واختلافاتٍ أخرى غير هذه
ضئيلة.
الدَّنا
DNA
المادةُ الوراثية التي تحملها الكروموزومات
هي الحمضُ النووي الديوكسي ريبوزي، أو الدنا،
وفيه تُخَزَّنُ المعلوماتُ الوراثية — وكما
يقول ليد أدلمان Led
Adleman فإن المعلومات
بالجرام الواحد من الدنا تُعادِل ما يحمله
مليون مليون قرصٍ مضغوط
CD.
وجزيء الدنا عبارة عن جديلتَين، كلٌّ تتألف
من تتابُعٍ
sequence
طويل لأربع قواعد
bases —
أو نوتيات
nucleotides
— هي الأدنين (أ) والثايمين (ث) والجوانين (ج)
والسيتوزين (س). والقاعدة أ على جديلةٍ
تُقابِلها القاعدة ث على الجديلة الرفيقة
المكملة (والعكس بالعكس)، أما القاعدة س
فتقابلها القاعدة ج (والعكس بالعكس). والجينُ
gene هو
امتدادٌ من أزواج القواعد على طول جديلَتَي
دنا الكروموزوم، ويُقاس طوله بعددها — فيُقال
إن طول هذا الجين مثلًا هو عشرةُ آلافٍ من
أزواج القواعد (أو ١٠٠٠٠ زق)، أو عشرة آلاف
نوتيدة. ومتوسط طول الجين البشري يبلغ نحو
ثلاثة آلاف زق، وإنْ كان هناك جين يصل طوله
٢٫٤ مليون زق (جين اسمه
dystrophin).
يُشفِّرُ الجينُ لبروتينٍ معيَّن، وذلك بأن
يُنْسَخ الجين بحامضٍ نووي آخر — هو الحامض
النووي الريبوزي، أو الرنا
RNA —
يخرج من نواة الخلية إلى السيتوبلازم لينفِّذ
التعليمات. تُوجَد التعليماتُ في الدنا في
صورة كودونات
codons
متتابعة والكودون عبارة عن ثلاث قواعد متتابعة
تشفِّر لحمضٍ أميني (وأول كودون في أي جين هو
أ س ج، الذي يشفِّر لحمض المثيونين الأميني) —
والبروتين سلسلة من الأحماض الأمينية، ترتيبها
يناظر ترتيبَ الكودونات بالدنا المُشفِّر له.
وفي وجود أربع نوتيدات فقط (أ، ث، ج، س) سيكون
لدينا ٦٤ كودونًا ممكنًا؛ ثلاثةٌ منها تحدِّد
نهايات الجينات، والباقي يشفِّر لأحماضٍ
أمينية. ولمَّا كان عدد الأحماض الأمينية
عشرين فقط، فقد يشفِّر للحمض الأميني الواحد
عددٌ من الكودونات؛ فحمض البرولين مثلًا
يُشَفِّر له أربعة كودونات هي س س ج، س س أ، س
س س، س س ث، أما حمض المثيونين فَيُشَفِّر له
كودونٌ واحد (أ س ج).
قد تحدث بالجين طفرةٌ نُقْطيَّة
point
mutation تتحول بها
قاعدةٌ إلى قاعدةٍ أخرى؛ فتتحول القاعدة أ
مثلًا إلى س أو ج أو ث. وقد يُغَيرُ هذا من
حمضٍ أميني بالبروتين الذي يُشفِّر له الجين،
وربما تسبَّبَ هذا في أن يصبحَ البروتينُ
الناتج مختلفًا.
وقد يحدُث أن تُحذف نوتيدة أو أن تُضَافَ
نوتيدة، ومن شأن مثل هذا الاقتضاب أو الإيلاج
أن يُحَوِّرَ كلَّ الكودونات التالية إذا وقع
في الدنا المشفِّر، فإذا كان الترتيب الأصلي
لمقطعٍ ما داخلَ جينٍ هو (أ س ج س س ث ج …)
وبه الكودونان أ س ج (يُشفِّر لحمض
الثرايونين) يليه الكودون س س ث (يُشفِّر لحمض
البرولين)، ثم اقتُضبَت أي حُذِفَت القاعدة أ
فسيكون الباقي س ج س س ث ج، ويصبح الكودونُ
الأول هنا هو س ج س (يُشفِّرُ للأرجنين)
والثاني هو س ث ج (يشفِّر للِّيوسين) وتتغير
أيضًا كل الكودونات التالية ليَنتُج بروتينٌ
جديد يختلف عن البروتين الأصلي. ومن الواضح أن
إضافةَ حرفٍ ستؤدي إلى نفس النتيجة. حرفٌ واحد
يُضافُ أو يُحْذَفُ يكفي لتغيير
البروتين!
سقَطُ الدَّنا
تُشكِّل الجيناتُ نسبةً ضئيلةً من دنا
الكائنات العليا، أما بقيةُ الدنا — الذي
يُسَمَّى سَقَط الدنا Junk
DNA — فلا تُعرف له حتى
الآن وظيفةٌ واضحة، وإن كانت نتائجُ الأبحاث
التي ظهرَت مؤخرًا (عام ٢٠٠٤م) تُشير، كما
سنرى إلى أن لهذا السَّقَط وظائفَ لم
تُكْتَشَف بعدُ. يبلغ طولُ الجينوم البشري نحو
٣١٠٠ مليون قاعدة، لا تُشكِّلُ الجيناتُ منه
أكْثَرَ من ٣٪. وقد يتخلَّل السَّقَطُ
الجيناتِ نفسها، في مناطقَ منها تُسمَّى
الإنترونات
introns
تمييزًا لها عن الدنا المُشَفِّر الذي
يُسَمَّى عندئذٍ بالإكسونات
exons.
يبلغُ طول جينوم نبات البصل ١٢ ضعفَ طولِ
الجينوم البشري، ويُفترضُ أن هذا يرجع إلى
زيادة نسبة السَّقَط فيه، أما جينوم السمكة
الكروية
Pufferfish،
الذي يتميز بخاصةٍ بقلَّة محتواه من سَقَط
الدنا، فلا يزيد طوله عن تُسعِ طول جينومنا،
لكنه يحمل تقريبًا نفسَ عدد جيناتنا. يبدو أن
نسبة سَقَط الدنا تختلف اختلافًا واسعًا بين
الأنواع. أما سببُ وجود كلِّ هذا السَّقَط،
فيعزوه البعض إلى أن ثمَّة جيناتٍ قد فقدَت
وظيفتَها وأصبحَت «جيناتٍ كاذبةً»، فَتَشَظَّت
وبقيَت عبئًا في الجينوم، بينما يرى آخرون أنه
مجرد مستودَعٍ من الدنا يمكن أن تنشأ منه
جيناتٌ أخرى مفيدة، فيما يعتقد البعضُ الآخر
أن له وظيفةً في التنامي أو في تعبير جيناتٍ
تُجاوره.
الدَّنا المَصون
يكتسبُ الدنا عادةً طفراتٍ عشوائيةً، فإذا
وقعَت هذه الطفراتُ في مناطقَ من الدنا ذاتِ
وظيفةٍ أساسيةٍ هامة، تسبَّبَت في موتِ
الأفراد. هذا يعني أن التتابعاتِ الدناويةَ
الأساسية للبقاء لا بد أن تبقَى محفوظةً بلا
تغيير، حتى بين الأنواع. وعلى هذا فعندما
يُقارِن الوراثيون جينوماتِ أنواعٍ مختلفة، ثم
يجدون مناطقَ من الدنا ثابتةً فيها، فإنهم
يعتبرون أن لهذه المناطق وظيفةً حيويةً
أساسية. من المستبعد بناءً على هذا أن يجدَ
العلماءُ مناطقَ مصونةً داخل سَقَط الدنا، إذا
كان هذا بالفعل بلا وظيفة، لكن الحقيقة هي أن
بعضَ العلماء قد وجَد في السَّقَط مثلَ هذه
المناطق المصونة من الدنا، واقترح أنها تؤثِّر
في تعبير الجينات المجاورة لها.
وتتابُع التيلومير (في طَرَفَي كل كروموزوم)
تتابعٌ مصونٌ للغاية في المجاميع المختلفة من
الكائنات. وعلى سبيل المثال فإن للفقريات
تتابعًا — هو: ث ث أ ج ج ج — يتكرر مراتٍ
ومراتٍ يصل تعدادُها إلى ما بين ٥٠٠ و٣٥٠٠ مرة
في الرئيسات (يبلغ طولُ التيلومير في
كروموزومات البشر نحو عشرة آلاف زق، أما في
الشمبانزي فيصل إلى ٢٣ ألف زق). يلاصقُ
التيلوميرَ، إلى الداخل مناطقُ بها مُكررات
دناوية أخرى، إن تكن غير مصونة تمامًا، فَبها
تبايناتٌ عديدة داخل النوع ذاته، لكن يسهُل
تمييزها في الأنواع الوثيقة القرابة.
نتائج مذهلة
لا يحدث كثيرًا أن يلهثَ الحاضرون في
اجتماعٍ علمي وهم يستمعون إلى عالِمٍ يتحدثُ
أمامهم على المنصة، لكنَّ هذا هو ما حدث يوم ٣
يونيو ٢٠٠٤م، عندما أعلن إدوارد روبين
E. Robin،
في جلسة اجتماعٍ عُقدت بمعمل سبرينج هاربور في
نيويورك، أن فريقَه العلمي قد اقتضَب مقاطعَ
ضخمةً من دنا جينوم الفأر، ولم يظهر أيُّ أثرٍ
ملحوظٍ على الفئران التي وُلِدَت «منقوصة
الدنا.» لم يكن مَن يتوقَّع هذه النتيجة؛
فالتتابعاتُ الدناويةُ المحذوفة كانت تَضُمُّ
مناطقَ مَصُونَةٌ يُظن أن لها وظائف
هامة.
كان العلماء يفترضون أن التتابعاتِ
المصُونَة لا بد أن تكونَ أساسًا في متن
الجينات التي تُشَفِّرُ للبروتينات، لكنهم
ذُهِلوا عند مقارنة جينومَي الفأر والإنسان؛
إذ وجدوا أن هناك بالفعل تتابعاتٍ كثيرةً
مصونةً داخل صحارى سَقَط الدنا لا تُشفِّر لأي
بروتين!
للتوصل إلى وظيفة مثل هذه المناطق المصونة
في سَقَط الدنا بالثدييات، قام فريقُ إدوارد
روبين في كاليفورنيا بحذف مقطعَين هائلَين من
هذا السَّقَط في الفئران، مقطعَين يحملان ١٠٠٠
تتابعٍ محفوظٍ شائعة بين البشر والفأر. كان
طولُ واحدٍ منهما يزيد على ٨٠٠ ألف قاعدة، أما
الآخر فيصل طوله إلى نحو ١٫٦ مليون قاعدة.
توقَّع الفريقُ أن تعانيَ الفئرانُ الناتجة من
مشاكلَ مختلفة في الحياة وفي البقاء.
لكن ما حدث هو أنَّ الفئران التي وُلدَت لم
تُظهر أي اختلافٍ واضحٍ عن الفئران الطبيعية
في كل الصفات التي قِيسَت؛ النمو، وظائف
الأيض، طول الحياة، التنامي. قال «روبين»:
«لقد ذُهلتُ فعلًا!» أَمَعقولٌ ألا تلعبَ
التتابعاتُ المصونةُ هذه أيَّ دور في النمو
والتنامي؟ إذن، فلأيِّ سَبَبٍ حُفظَت؟ ربما
كانت هذه التتابعاتُ تخدمُ غرضًا مراوغًا لم
تتطرق إليه الاختباراتُ التي أجراها الفريقُ
على الفئران المنقوصة الدنا؟ أم تُرى أن
تتابعاتٍ أخرى، بمناطقَ أخرى من الجينوم، قد
قامت على الفور بتأدية وظيفة الجزء المُقتضَبِ
من الدنا، فلم يظهر للحذف أثر؟
مشروع الجينوم الشمبي
في أغسطس عام ٢٠٠٠م، طالَب لفيفٌ من علماء
الوراثية الأمريكيين بإقامة مشروعٍ لجينوم
الشمبانزي. وفي يناير ٢٠٠٣م بدأ المشروعُ
بالفعل استخدامَ دنا ستة أفراد، وكان مشروع
الجينوم البشري آنذاك على وشك الانتهاء
(انتهَى في أبريل ٢٠٠٣م). كان من أهم
المبرِّرات التي قُدِّمَت لإقامة المشروع، أن
مقارنةَ التفاصيلِ الجُزَيئية لجينوم
الشمبانزي، بمثيلاتها البشرية قد تُساعِد في
الوصول إلى علاجاتٍ بشرية لأمراضٍ خطيرة
للغاية؛ فالشمبانزي مثلًا لا يُصاب بالملاريا
التي تقتل ملايين البشر، وهو لا يُصاب بمرضِ
الإيدز. وعلى الرغم من أنه يُشاركُنا في جينٍ
يجعلنا عرضةً للإصابة بمرض الألزهايمر، فإنه
لا يُصاب به؛ لأن جينه يُنتج بروتينًا مختلفًا
بعضَ الشيء. ثم إن نسبةَ إصابته بسرطانات
القولون والمعدة والثدي والبروستاتا منخفضةٌ
كثيرًا عنها في البشر.
وفي ١٠ ديسمبر ٢٠٠٣م، أُعلن عن الانتهاء من
خَرْطَنَة
mapping
مسوَّدة جينوم الشمبانزي — الخريطة الكاملة مع
بعض فجواتٍ وأخطاءٍ قليلة — ونُشر ذلك في عدد
١٢ ديسمبر من مجلة «ساينس.» كانت تكاليفُ
السلسلة قد بلغَت نحو ٢٠ مليون دولار. وفي ٢٧
مايو ٢٠٠٤م ظهر بمجلة «نيتشر» تقريرٌ لخمسةٍ
وأربعين عالِمًا يُعْلِنُ الانتهاء من سلسلة
sequencing
نوتيدات كروموزوم ٢٢ الشمبي (ويبلغ عددها نحو
٣٣٫٣ مليون قاعدة) بدقة بلغَت ٩٩٫٩٩٨٪، نفس
الدقة التي فُحصَت بها كروموزومات الجينوم
البشري. وهذا الكروموزومُ الشمبي هو النظير
للكروموزوم ٢١ البشري — الكروموزوم المسئول عن
متلازمة داون أو الطفل المغولي (التي تظهر في
الإنسان وفي الشمب) ومرض الألزهايمر،
واللوكيميا الحادة. والمقارنة بين
الكروموزومَين ستساعد في كشفِ البعضِ من
الطفرات التي أدَّت إلى الفروق بين النوعَين،
والعوامل المختلفة التي لعبَت دورًا رئيسيًّا
في تطوُّر الرئيسات.
نتائج التشريح الجزيئي لجينوم
الشمبانزي
تقول النتائجُ التي ظَهَرَت أن درجةَ تطابُق
تتابعِ النوتيدات في جينومَي الشمب والإنسان
تصل إلى ٩٨٫٧٪. قد يُشير هذا إلى أن عددًا
محدودًا فقط من الجينات هو الذي يُحَدِّدُ
جوهر «البشرية.» رصَد العلماء ٧٦٤٥ جينًا
مشتركًا بين الإنسان والشمب والفأر، وبحثوا
بينها عن الجينات التي كابدَت التغيُّر،
ومَيَّزَت هذا عن ذاك، فاتضح أن تغيُّراتٍ
سريعةً قد وقعَت في ١٥٤٧ جينًا بشريًّا (ومن
ثَم في البروتينات التي تُنتِجُها) وفي ١٥٣٤
جينًا شمبيًّا — وذلك بمقارنة هذَين النوعَين
بالفأر، الأدنى مرتبةً على سُلَّم التطور. يرى
البيولوجيون إذن أن الانتخاب الطبيعي قد
حَوَّر في البشر ١٥٤٧ جينًا أضفَت علينا صفةَ
البَشَرِيَّة، حُوِّرَت لتتلاءمَ مع البيئة
التي نحيا بها. وقد قَدَّر ديريك وايلدمان
D. E.
Wildeman وزملاؤه أن عدد
الجينات التي مَيَّزَتْنَا يتراوح ما بين ٢٨٠٠
و٤٠٠٠، وهذه تختلف جوهريًّا عن نظيراتها في
الشمبانزي. والبعضُ من هذه الجينات لا بد وأن
قد تَخَصَّص في تنظيم عمل جيناتٍ غيرها؛
فالجينات التنظيمية تعمل في فتحِ أو إغلاقِ
البعضِ غيرَها من الجينات؛ ومن ثَم تُشجِّعُ
أو تُثَبِّطُ إنتاج بروتينات هذه الجينات
الأخرى. وتغييرُ جينٍ مُنَظِّم واحد قد
يُغيِّر، ليس فقط من إنتاج بروتيناتٍ عديدة،
بل وأيضًا من توقيتِ إنتاجِها وموقعِه في
الجسم.
فَحَص روي بريتِّين Roy J.
Britten ٧٧٩ ألف نوتيدة
في الشمبانزي وفي الإنسان، وأعلن في تقرير
للأكاديمية الأمريكية للعلوم أن ١٫٤٪ من
النوتيدات قد استُبدلَت. جاء هذا التقريرُ
نتيجةَ فَحْصِ الطفراتِ النُّقْطِيَّةِ التي
تؤثِّر في نوتيدةٍ واحدةٍ فقط. هناك بالطبع
طفراتُ حذفٍ أو إضافة (إنديلات
indels)
تحدُث في مقاطع طولها ٢–٤ نوتيدات، بل وهناك
قلَّةٌ يَصِل طول الإنديل منها إلى أكثر من
١٠٠٠ نوتيدة. وضَع بريتِّين هذا الأمر في
اعتباره، فجاء تقديرُه للتطابُق بين
الجينومَين: ٩٥٪. واختلافٌ قَدْرُهُ ٥٪ من
الدنا يعني في الحقيقة فارقًا بين النوعَين
يصل إلى ١٥٠ مليون نوتيدة!
نتائج كروموزوم ٢٢ الشمبي
وعند مقارنة نتائج تحليل كروموزوم ٢٢
الشمبي، الذي تمَّتْ سَلْسَلَتُه النهائية،
بنظيراتها على الكروموزوم ٢١ البشري، اتضح
وجودُ فروقٍ قدرها ١٫٤٤ ناتجة عن استبدالات
لقواعد دنا مفرَدة. هذا الفارق الضئيل كان
كافيًا لتغيير تتابع الأحماض الأمينية في ٨٣٪
من البروتينات التي يُشفِّر لها ٢٣١ جينًا
وُجدَت مشتركةً بين الكروموزومَين! والحقُّ
أنَّ اختلافَ البروتيناتِ بين النوعَين هنا
كان في حفْنةٍ ضئيلة من الأحماض الأمينية
داخلها؛ ومن ثَم فمن المفترض أن تَظَلَّ
وظائفُها متشابهةً للغاية.
اتضح أيضًا أن هناك نحو ٦٨٠٠ منطقة في الدنا
— لا يزيد طولُ معظمها عن بضعِ قواعد، وإنْ
وَصَلَ طولُ البعض منها إلى ٥٤٠٠ نوتيدة — قد
حَدَثَ بها إيلاج أو اقتضاب، وكانت هذه كافيةً
لتسبِّب فروقًا ضخمةً في تركيب أكثر من ٢٠٪ من
البروتينات، وهذا اختلافٌ يزيد كثيرًا عما كان
متوقعًا، ثم إن البحث في وقت فتح الجينات
ودرجة فتحها قد أوضح أن ٢٠٪ من الجينات قد
بَيَّنَت «فروقًا معنوية في نمط
نشاطها.»
كيف نُحَدِّدُ إذن درجة التشابه بين
الأنواع؟ هذا أمرٌ قد يكون ذاتيًّا، لا
موضوعيًّا. إذا نظرنا إلى شدة التشابه بين
نوتيدات الدنا فسنجد تشابهًا قد يصل إلى ٩٨٪
بين جينومَي البشر والشمبانزي، فإذا أخذنا في
اعتبارنا المولجات والاقتضابات (الإنديلات)
تَحَوَّلَت درجة التشابه إلى ٩٥٪! أما على
مستوى الجينات فقد اتضح أن كل الجينات
المعروفة الموجودة على الكروموزوم ٢١ البشري
موجودةٌ أيضًا على الكروموزوم ٢٢ الشمبي، لكن
كان ثمَّة اختلافاتٌ جُزيئية في الكثير منها.
وسنعرض الآن نبذةً عن بعضِ الجيناتِ الهامة
المشتركة، والمختلفة في نفس الوقت، في جينومَي
البشر والشمبانزي.
جين للُّغة
عرف المجتمع العلمي عن عائلةٍ (أُطلِق عليها
اسم K. E.)
منذ نحو عام ١٩٩٠م، كان نصفُ أفرادها، وعلى
مدى ثلاثة أجيال، يُعانون من مشاكلَ أوضحُها
الصعوبة في الكلام، حتى لَيُصبح حديثهم غيرَ
مفهوم، فيتطلَّب الأمر تدريبَهم وهم أطفال على
لغة الإشارة. ظهر من البداية أن هذه حالةٌ
وراثية، وأنها صفةٌ سائدة أوتوزومية (أي غير
مرتبطة بكروموزوم الجنس س
X) يُعاني
فيها المصابون من قصورٍ في إنشاء الجُمَل وعدم
التمكُّن من تفهُّم بعض نواحي قواعد اللغة،
ومن عدم القدرة على تشكيل كلامٍ مفهوم،
وقُصُورٍ في القدرة على تحريك الفَك الأسفل
والفم، لا سيما الشفة العليا واللسان، بجانب
انخفاض مُعامل الذكاء (المتوسط = ٨٢
نقطة).
ظهر أولُ بحثٍ عن هذه العائلة في مجلة
«نيتشر» في ٤ أكتوبر ٢٠٠١م، واتضح أن الجينَ
المسئول (وقد أُطلِق عليه اسم
FOXP2)
موجودٌ على الكروموزوم السابع، وأن طفرةً بهذا
الجين تغيَّرَت به نوتيدةٌ واحدة فقط — من بين
الخمسمائة نوتيدة التي تؤلف الجين — هي
المسئولة عن المرض بعائلة K.
E.؛ إذْ فقَد بها هذا
الجين قدرتَه على تنشيط التتابع الطبيعي
للجينات اللازمة للتنامي المبكِّر للمخ، مما
تسبَّب في تغيرٍ غيرِ طبيعي في عُقَدٍ عصبيةٍ
بالمُخ تختصُّ بالحركة (ومن هنا صعوبة تحريك
الشفتَين واللسان).
وقد اتضح أن هذا الجين جينٌ تنظيمي مَصُون،
يشفِّر لبروتين يتألف من ٧١٥ حمضًا أمينيًّا،
وتُوجَد منه صيغٌ مختلفة في الشمبانزي
والغوريلا والأورانج يوتان وقرد الريزوس
والفأر — وإنْ كُنَّا لا نفهم له وظيفةً لدى
هذه الحيوانات جميعًا. تُبيِّن الصيغةُ
البشرية للجين دلائلَ على تغيرٍ تطوري متسارع
حدث خلال المائتي ألف سنة الأخيرة، اكتسب به
الجينُ وظيفةً جديدةً ساعدَت في إضفاء مَلَكَة
الكلام على البشر؛ فثمَّة تحورٌ طفيفٌ بهذا
الجين لم يُعثر عليه في أي ثدييٍّ آخر — يتحول
فيه حمض ثريونين في الصِّيغ غير البشرية إلى
حمض أسباراجين — قد تَسَبَّب في أن يُطَوِّرَ
الإنسانُ بِنْيَة الوجه والفَك والقدرةَ على
التحريك الرهيف للفم والحنجرة، لتصبح
إمكاناتُنا المتفردةُ على الكلام واللغة أمرًا
ممكنًا.
هذا الجين نَشِطٌ جدًّا، وهو يعمل في الكبد
والرئة والمخ، وله على الأغلب مهامُّ أخرى في
الجسم، لكنه يبدأ نشاطَه مبكرًا جدًّا في مخ
الجنين أثناء تناميه في رحم الأم، ويبلغ
نشاطُه غايتَه في مناطق المخ التي تتحكَّم في
الحركة.
يُشكِّل هذا الجين، على ما يبدو، بدايةَ
وضعِ الأساس الوراثي للُّغَةِ البشرية. هو ليس
جينَ «اللُّغة»، إنما هو جينٌ «للغة.» واللغة
صفةٌ يتفرد بها البشر. من الممكن أن يدرَّب
الشمبانزي على الاتصال بالآخرين باستخدام
مجموعةٍ معقدةٍ من الإشارات، لكنه لا يستطيع
أن يقومَ بحركات الوجه المطلوبة. باللغة
تَمَكَّنَ الإنسانُ من نقل كمياتٍ هائلة من
المعلومات بين الأجيال؛ ومن ثَم فقد كانت قوةً
دافعةً هامة في انتشار الإنسان على وجه الأرض
وفي قيام الحضارة.
جين للسَّمع
ربما كانت الجيناتُ التي تُمَكِّنُ الإنسانَ
من تفهُّم اللغة لا تعمل في المخ وحده، وإنما
تعمل أيضًا على حاسة السَّمْع. جاءت الإشارةُ
الواضحة على هذا من جينٍ أُطلِق عليه اسم ألفا
تكتورين alpha
tectorin (ويقع على
الكروموزوم ١١ البشري)، يُشفِّر لبروتينٍ
غامضٍ في الغشاء التكتوري بالأذن الداخلية
للإنسان، وتُؤدي طفرة (سائدة) فيه إلى إحدى
صور الصَّمَم. وهذا الجين البشري (وهو أحد
ثلاثة، من بين ٢١ جينًا مسئولة عن السمع،
كابدَت تغيراتٍ متسارعة) يختلف كثيرًا عن
نظيره في الشمبانزي، ويُظَن أن الصورة البشرية
قد تطوَّرَت على الأغلب مع جينات الكلام،
لتجعلَ حاسةَ السمع في الإنسان أكثر حدةً
وأكثر دقَّة — فهذا أمرٌ يصبح ضروريًّا
لتفهُّم اللغة المعقَّدة المنطوقة — حتى ليرى
البعضُ أن صعوبةَ تدريب حيوانات الشمبانزي على
تفهُّم لغتنا إنما ترجع إلى أن حاسة السَّمْع
لديها لا ترتقي إلى المرتبة التي نتمتع بها
نحن البشر.
جين لهذا السُّكَّر اللعين
من بين الجينات التي تختلف بين البشر
والشمبانزي، هناك الجين
CMAH الذي
يُشَفِّرُ لإنزيم يصنع سُكَّرًا يغلِّف أسطح
خلايا الجسم — سُكَّرًا يُطلق عليه اسم
Neu5Gc.
يُوجَد هذا السكر على أغشية خلايا جسم
الشمبانزي وغيره من الثدييات التي دُرسَت،
لكنه غير موجود في البشر؛ ففينا يصبح هذا
الجين بلا عمل؛ ثمَّة مقطعٌ طويل من سَقَط
الدنا قد حلَّ محلَّ جزءٍ من الجين فأصبح بلا
عمل. يتألف البروتين الذي يُشفِّر له جينُ
الشمبانزي من ٥٨٠ حمضًا أمينيًّا، أما بروتين
الجين البشري فمن ٧٢ حمضًا فقط. هذه الصيغة من
الجين العديمة الجدوى تُوجَد الآن في كُلِّ
إنسانٍ على سطح الأرض، ولا تُوجَد في أي
ثدييٍّ آخر.
يُوجَد هذا السكَّر إذن على أغشية كل خلايا
جسم الشمبانزي وغيره من الثدييات، لكن في
مخاخِ نفس هذه الحيوانات تعمل جيناتٌ أخرى
تُبطئ من إنتاج هذا السكر، فلا نجد منه إلا
القليل جدًّا على أسطح الخلايا العصبية
(النيورونات)؛ الأمر الذي قد يشير إلى أن لهذا
السكَّر أثرًا ما جانبيًّا ضارًّا على خلايا
المخ — لا نعرفه حتى الآن. وعلى هذا يُمكِن
القول إن الإنسانَ قد حظِيَ بنعمةٍ غاليةٍ
بتوقُّف الجين
CMAH هذا
عن العمل بخلاياه، فربما كان هذا من بين أسباب
استمرار تطوُّر مخه إلى الحد الذي جعله يتميز
عن كل مخلوقات الله.
جينات أخرى مشتركة
هناك فروقٌ أخرى بين البشر والشمب فيما قد
يصل إلى ٨٠ جينًا تَخْتَصُّ بإعادة تدوير
الأحماض الأمينية عند هضم البروتينات، وهذه
الفروقُ قد تعكس ما حدث من تَغَيُّر في الطعام
عندما تَحَوَّلَ الأسلافُ من طعامٍ نَبَاتِي
أساسًا إلى أكل اللحوم.
ثمَّة اختلافاتٌ أخرى قد حدثَت في الجينات
التي تعمل على حاسة الشم، الحاسة التي تختلف
بوضوحٍ بين البشر والشمبانزي؛ فالكثير من
الجينات الخمسين المهمة المرتبطة بالشم، التي
تشفِّر لمُستقبِلات في الأنف، تبدو في البشَر
وقد توقفَت تمامًا عن العمل، وأصبحَت «جينات
كاذبةً»، ربما لتعكس تضاؤل أهمية الشمِّ في
أسلوب حياتنا مقارنةً بالشمب. في نفس الوقت،
هناك من بين هذه الجينات الخاصة بالشم جيناتٌ
قد تطوَّرَت وبسرعة في البشر، وتَضُمُّ هذه
الجيناتِ اللازمةَ للعثور على الطعام وتمييز
طبيعته وطزاجته (وربما أيضًا لاختيار الرفيق).
صحيحٌ أن حاسة الشم قد تدهورَت كثيرًا في
البشر، لكنَّ اللهَ قد عوَّضَهم عنها برؤيةٍ
للألوان أفضل، ومقدرةٍ أفضل على التواصل مع
الآخرين عن طريق الوجه بديلًا عن
الرائحة.
هناك جيناتٌ أخرى تختص بتنامي المخ، من
بينها جينٌ هام أُطلِق عليه اسم
SEMA3B
يساعد في توجيه المحاور العصبية
axons
النامية إلى المناطق الصحيحة من الجسم،
والفروق في الصيغة البشرية لهذا الجين مقارنةً
بالصيغة الشمبيَّة قد تفسِّر بعضًا من
الاختلافات في الشبكة العصبية بين البشر
والشمبانزي.
ولقد حدَّد الباحثون أيضًا هُويَّةَ جينَين،
هما NCAM2
وGRIK1،
لهما علاقةٌ بوظائف الأعصاب، وكانت الصيغة
البشرية لكلٍّ منهما تحمل مقاطعَ طويلةً من
الدنا لا تُوجَد في الشمبانزي، يصل طولها في
الجين الأول إلى ١٠٠٠٠ نوتيدة، وفي الجين
الثاني إلى ٤٠٠٠ نوتيدة.
عن الكروموزوم البشري رقم ٢
مثلما
وجد الباحثون كلَّ هذه الفروق بين البشر
والشمبانزي، فقد وجدوا أيضًا طفراتٍ مختلفةً
في هيكل الكروموزومات ذاتها. للشمبانزي جينوم
يضُم أربعةً وعشرين زوجًا من الكروموزومات،
ويحمل هذا الجينوم — تقريبًا — نفس عدد
النوتيدات الموجودة بالجينوم البشري المكوَّن
من ثلاثة وعشرين زوجًا. اكتشف العلماء بمقارنة
أنماط التشريط على الكروموزومات في كلا
النوعَين أن الكروموزوم رقم ٢ البشري قد يكون
ناتجًا عن «انتقال» اندمج فيه كروموزومان
صغيران من جينوم الشمبانزي (انظر الشكل
٢)، بمعنى أن اختلافنا عن
الشمب في عدد الكروموزومات لم ينشأ عن فَقْدنا
لكروموزوم، وإنما عن التحام اثنين من
كروموزومات الشمب ليظهرا ككروموزومٍ واحد،
فإذا كن هذا صحيحًا، فإن معناه أن تيلوميرَيْ
هذَين الكروموزومَين الصغيرَين لا بد وأن قد
اندمجا ليمكثا في وسط الكروموزوم البشري.
والتيلومير هنا يتكوَّن من عددٍ كبيرٍ من
مكرراتٍ كلٌّ منها، كما ذكرنا، مؤلَّف من ست
نوتيدات هي ث ث أ ج ج ج، ووجودُ هذه المكررات
بالمئات في وسط الكروموزوم — أي كروموزوم، في
أي كائن — هو أمرٌ مستحيل؛ فهي لا تُوجَد إلا
في أطراف الكروموزومات وتلاصقها، وقبلَها
مباشرة، تُوجَد بالطبع المكرراتُ المصاحبةُ
للتيلومير.
بالفحص الجُزَيئي لتتابعات النوتيدات بوسط
الكروموزوم ٢ البشري اتضح أنه يحمل:
[تتابع ما قبل التيلومير – تتابع التيلومير
– تتابع التيلومير مقلوبًا – تتابع ما قبل
التيلومير مقلوبًا].
بهذا الترتيب! ومقلوبُ تتابعِ المكررِ هنا
يعني س س س ث أ أ؛ فهذه هي القواعدُ الستُّ
المكملةُ للمكرر الأصلي (ث ث أ ج ج ج)
مقلوبًا. ما الذي جاء بهذه المكررات في وسط
الكروموزوم؟ سوى أن يكون بالفعل ناتجًا عن دمج
كروموزومَين؟
ثم إن سنتروميرَ الكروموزوم ٢ البشري يُوجَد
في نفس مكانه بالكروموزوم ٢ ق الشمبي (الأقصر)
— كذا يقول تَشابُه التتابع — كما أن تتابُعَ
النوتيدات في الذراع الطويلة للكروموزوم ٢
البشري تحمل البقايا المؤكدة لنوتيدات
السنترومير الأصلي للكروموزوم ٢ط الشمبي
(الأطول) (وهو بالطبع لا يعمل كسنترومير
بالكروموزوم البشري).
يبدو أن الشواهدَ على أن الكروموزوم الثاني
البشري قد جاء عن دمجِ كروموزومَي الشمبانزي
شواهدُ دامغةٌ فعلًا.
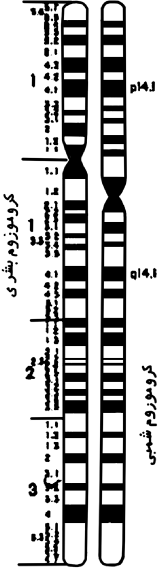
طفرة انقلاب في الكروموزوم البشري رقم
٥
إذا قَارنَّا بين تشريط الكروموزوم البشري
الخامس والكروموزوم الشمبي النظير (انظر الشكل
٣) فسنُلاحظ أن
الكروموزومَين متماثلان، ويتطابقان في
التشريط، وفي موقع السنترومير، إذا قَلَبْنَا
المنطقة في الكروموزوم الشمبي ما بين
p14.1
و
q14.1؛
الأمر الذي يشير إلى أن الكروموزوم البشري قد
جاء عن طفرة انقلابٍ في هذه المنطقة من
الكروموزوم الشمبي.
ما بين الدَّنا
(DNA)
والدِّين
ثمَّة شيءٌ من التوتُّر يَسُود دائمًا
العلاقةَ بين رجالِ العلم ورجالِ الدين.
العلمُ يُقدِّم نتائجه، وتفسيرُها عادةً ما
يكون السببَ في هذا التوتُّر. العلمُ بالضرورة
يقود إلى الإيمان، والدينُ بالتأكيد يدعو إلى
العلم. ومن الصعب أن نجد تبريرًا لهذا
التوتُّر إلا إذا كان منشَؤه الاختلاف في
وجهات النظر عند تفسير نتائج العلم، أو عند
تفسير دلالات الألفاظ في الكتب المقدَّسة.
عندما نشر جاليليو كتابه «حوار عن نُظُم
العالَم الفسيح»، سنة ١٦٣٢م، مُعضِّدًا فكرةَ
كوبرنيق بأن الأرض هي التي تدور حول الشمس —
لا العكس — حُوكم أمام محكمة تفتيشٍ حكَمَت
عليه بالسجن في ٢٢ يونيو ١٦٣٣م. كانت آراءُ
جاليليو تُعارِض آراءَ بطليموس وأرسطو
الضَّحلة، ولم تكن تُعارِضُ الإنجيل، سوى أن
الكنيسةَ كانت قد اعتَبرَت آراءَ هذَين من
صُلب الدين، وأن الخروج عليها كُفْر. وبعد ٣٥٩
سنة، وفي ٣١ أكتوبر ١٩٩٢م، ألغى البابا جون
بول الثاني الحُكْم، بعد أن اتضح له أن
الكنيسةَ قد أخطأَت.
أحسَّ الناسُ عندما ظهر كتابُ جاليليو بأن
الأرض قد فقَدت مكانها الأسمى، ولم تعُد هي
مركز الكون. ولمَّا نشَر داروين كتابَه «أصل
الأنواع» عن نظرية التطور، أُصيب الناس
بالفزع، هذه المرة لأنهم أحسُّوا بأن الإنسان
يفقد بها مكانتَه الرفيعة كسيِّدٍ للكون.
يُروى آنئذٍ أنَّ زوجةَ أُسْقُف ورشستر عندما
سمعَت عن نظرية داروين صرخَت قائلةً: «يا
فَضيحتي! نحن من نسل القرود؟ آمُل ألَّا يكونَ
هذا صحيحًا، فإذا كان صحيحًا فأرجو من الله
ألا يعرفَ به أحد.» وقفَت الكنيسةُ صراحةً ضد
النظرية، وربما كان أول صدامٍ علني هو ما حدَث
بين ديلبرفورس وهكسلي بعد ستة أشهر من صدور
«أصل الأنواع.»
يقول مُعارِضو نظرية التطور من رجال الدين —
ورجال العلم أيضًا — إنه إذا كانت الأبحاثُ قد
أثبتَت صحةَ ما جاء به جاليليو، فإن نظرية
التطور لم تثبُت، وإن أحدًا لم يلحَظ التطور،
وليس ثَمَّ أحافيرُ انتقالية تُمثِّل كُلَّ
الصُّور التي مَرَّت بها الكائنات الحية عبْر
تطوُّرها؛ فالسجل الحَفْرِيُّ لا يبيِّن
عمليةَ التطور، إنما يكشفُ عن الظهور الفجائي
للصور الجديدة من الحياة. ثم إن الوضعَ مع
نظرية التطوُّر مختلف؛ فالإنجيل قد ذكَر
صراحةً أن الله قد خلَق الكونَ كلَّه في ستة
أيام — والإنسان به — ورجالُ التطور يدَّعون
أن الإنسانَ قد تَطوَّر عبْر ملايين السنين.
والمعلوماتُ المُضَمَّنَةُ في الدنا ليست من
صُنع الدنا؛ فمِنْ أين جاءت؟ مَن يكون
مؤلِّفها؟ ثم ماذا يحدُث لو اكتشَفنا بعضَ
الجينات التي تُضفي علينا صفات «البشرية»، ثم
رأى بعضُ العلماء أن يُولجوها في الشمبانزي.
مَن يا تُرى سيتحملُ نتيجةَ مثل هذا التهجين
الدناوي؟ ومَن يا تُرى يتحمل نتيجةَ أن يحاولَ
العلماءُ هندسةَ بشرٍ أكثر «بشرية»، يجمَعون
بهم «أفضل» صِيَغِ الجينات التي نتفرَّد بها؟
ثم إذا كان الصحيح أن ١٨ زوجًا من
الكروموزومات تكاد تكونُ متطابقةً في البشر
والشمبانزي فإن الكروموزومات ٤ و٩ و١٢ تبيِّن
شواهد على أنه قد «أُعيد تشكيلها»، نعني أن
الجينات الواسمات
markers
على هذه الكروموزومات الثلاثة ليست بنفس
الترتيب في النوعَين. وبدلًا من «إعادة
التشكيل» التي يقترحُها التطوريُّون، لماذا لا
نقول إنها فروقٌ جوهرية بسبب «الخَلْق
المُنفصِل» الذي يقول به الخَلْقَويُّون؟ هم
يؤكِّدون أن القرنَ العشرين قد شهد سقوطَ
اثنَين من «الثلاثة الكبار»، الذين كان لهم
أكبر الأثر في تشكيل الحضارة الغربية؛ سيجموند
فرويد S.
Freud وكارل ماركس
K. Marx،
وأن القرنَ الحاديَ والعشرين سيشهد سقوطَ
الثالث؛ تشارلس داروين.
لكن «فرانسيس كولينز» F.
Collins رئيس مشروع
الجينوم البشري يقول: «إنني أرى أن الله
بحكمته قد استخدم التطور مُخَطَّطًا للخَلْق،
ولا أعرف سببًا في أن تكونَ فكرتي هذه سخيفة.»
ثمَّة دراسةٌ مثيرة قام بها الصديق الدكتور
«حسن عطية» في أحد كتبه؛ فقد حاولَ تفسيرَ
معاني الكلمات: «الناس، البشر، الإنس،
الإنسان» — كما جاءت بالقرآن الكريم — ورأى
باجتهاداته أنها لا تعني نفس الشيء، وأن
تفسيراته لها قد تحتوي اجتهادات العلماء بشأن
نشأة الجنس البشري. أما «البابا جون بول
الثاني»، فقد وجَّه رسالةً في ٢٣ أكتوبر
١٩٩٦م، يقول فيها إن النتائج التي تجمَّعَت
عبْر الخمسين سنةً الماضية قد أدَّت إلى
الاعتراف بأن نظرية التطور هي أكثرُ من مجرد
فَرْض، وأن الخَلْقَويةَ
creationism
والتطوُّر يمكن أن يعملا سويًّا دون تضارب،
طالما أكدنا أن اللهَ لا سواه هو مَن يَنفُخ
الروحَ في البشر. والواقع أن التَّطوريين
والخَلْقويين سويًّا لا يعملون في العلم
التجريبي عندما يُعالجون قضيةَ نشأة الحياة؛
فهذه لا يمكن أن تُكَرَّر، أو أن تُوضَعَ في
أنبوبة اختبارٍ لتُفحَص. إن ما يقومون به
جميعًا هو تمارينُ جدليةٌ يحاول فيها كل فريقٍ
أن يطرح نظريةً حول الماضي ترتكز على بياناتٍ
تجريبيةٍ نلاحظها اليوم. والنظرية التي
ستُقدَّم ستكون، أولًا وأخيرًا، مجرد هيكلٍ
فلسفي لتفسير آخرِ البيانات التي جُمعَت
«فالحقيقةُ بنتُ الزمن»، كما يقول المثلُ
اللاتيني القديم!
النتائجُ العلمية التي عُرِضَت هنا عن
الجينوم البشري وجينوم الشمبانزي، صمَّمها
وجمَعها ونشَرها وفسَّرها مَن يُعضِّدون
التطور. كُلُّ تفسيراتهم تقول إن البيئة
والانتخاب الطبيعي كانا من وراء التباينات في
الدنا بين النوعَين. وجدوا أن تشابُه تتابعات
الدنا بينهما أوثَقُ من أن يكون مجرد صدفة،
فقالوا إنهما قد انشَعَبا عن أصلٍ مشترك. ثم
إنهم قدَّموا إثباتَهم العجيبَ على أن
كروموزومًا بشريًّا قد جاء عن اندماج
كروموزومَين شمبيَّين، وأن كروموزوماتٍ بشريةً
أخرى قد جاءت عن انقلابات داخل كروموزومات
شمبية، لكنَّ معنى التشابه وقَدْرَه يختلفان
حتى بين العلماء. لقد وفَّر تتابعُ الدنا لهم
معيارًا جديدًا يقيسون به التشابه بين كائناتٍ
كان من المستحيل قَبْلًا أن تُجرى بينها
مقارنةٌ ذات معنًى، لكن المعيار على ما يبدو
لا يزال يحتاج إلى ضبط. التشابه بين دَنَانَا
ودَنَا الشمبانزي كما رأينا يبلغ نحو ٩٨٫٧،
ورغم ذلك فحَجْم مُخِّنا يبلغ مثلَين أو ثلاثة
أمثال حجم مُخ الشمبانزي، كما نتميَّز عنه
بالقدَمانية
bipedalism؛
أي المشي على قدمَين والثقافة المتطورة
وبالحجم الأصغر للأسنان الخلفية، فإذا نظرنا
إلى مدى التشابه بين دنانا ودنا كائنات أخرى،
رأينا العجب؛ فهو مع الغوريلا ٩٨٫٤٪، ومع القط
٩٧٪ (ودنا الكروموزوم
A1 في
القط مؤلَّف من أجزاء من الكروموزومَين ٥ و١٣
البشريَّين)، ومع الكلب ٩٥٪، ومع الحصان ٨٠٪
(وكل دنا الكروموزوم البشري الثامن موجودٌ
بالكروموزوم التاسع للحصان)، ومع الفأر ٨٥٪،
ومع نبات الذُّرة ٦٧٪، ومع نبات الموز ٦٠٪،
فهل مَن يقول إن نبات الموز نصفُ بشر؟! أو إن
الذُّرةَ ثُلُثَا بشر؟! ليس على أيٍّ منا أن
يبحث عن الشمبانزي بداخله!
لكن، ماذا يعني هذا التشابُه حقًّا؟ وكيف لا
نتوقَّعه؟ لقد خلَق الله الكائنات جميعًا ولها
نفس المادة الوراثية، حتى ليُمكِن بالهندسة
الوراثية أن ننقلَ الدنا بين الكائنات جميعًا،
نحن وكل حي أقارب في الدنا. كذا حي كانت
مشيئتُه، «وما كانت مشيئتُه بلا معنًى»، كما
يقول الشاعر «إيليَّا أبو ماضي». لو شاء لجعَل
لكُلٍّ مادتَه الخاصة من المستحيل ألَّا تكونَ
بيننا وبين كلِّ الأحياء صلَة. كلنا من عجينةٍ
واحدة، أو قُلْ من «طينة» واحدة، اسمها الدنا،
يختلف مقدارُها، ترتيبُها، تجميعُها،
معمارُها، فتختلف الكائنات، وتتشابه. الله لم
يميِّزنا نحن بمادةٍ وراثيةٍ تَخصُّنا وحدَنا
ولا يشترك فيها معنا الشمبانزي أو غيره.
والبحث في الدنا المقارن يُوسِّع قدراتنا،
وخيالنا، وفَلْسَفَتَنَا، وتَفَهُّمَنَا
للحياة، ويُفسِح المجال أمام اكتشاف علاجاتٍ
جديدة للأمراض لم تكن تخطُر لنا قَبْلًا على
بال!
ومثلما يقول «إيليا أبو ماضي»: «أَرَادَ
اللهُ أَنْ نَعْشَقَ لمَّا أَوْجَدَ
الحُسْنَا»، فربما كان لنا أيضًا أن نقول:
«أَرَادَ اللهُ أَنْ نَعْرِفَ لمَّا أَوْجَدَ
العَقْلَا»! فلو أراد ألا نعرفَ لسوَّانا بلا
عقلٍ يُفكِّر!